

Landscape genetics has grown feverishly since its first formal definition in 2003 (Manel et al). The beauty of combining genetic, environmental, and spatial variation to answer biological questions sure is alluring, and the quest for improving the methodology of landscape genetics has been a reoccurring theme.
Entire issues are devoted to the subject in Evolution, Molecular Ecology, and Landscape Ecology. It is a broad field with a lot to talk about, and that comes with quite a few reviews over the years. Almost every review paper includes summaries of the topic and a dedicated section of future issues. Tracking the disparities between the “past” and “future” between review articles can give you an informative trajectory of a field: what problems are solved, what problems linger on, and what might not get solved any time soon.

Data return from SCOPUS using the search terms (landscape AND genetics). You can go play with the data yourself here.
In a (perhaps foolish) effort to distill some of the major issues down to a coffee break read, I’ve chosen some of the most influential reviews on landscape genetics (or landscape genomics) and broke down the evolution of the following components: the questions, the data, and the analyses.
The Questions
In the initial paper introducing the term “Landscape Genetics”, Manel et al (2003) recognized the need for a discipline that exists at a smaller spatial scale than biogeography and more contemporary temporal scale than phylogeography. In 2007 and again in 2010, Storfer et al looked back on the previous six years and revealed that most studies focused solely on detecting barriers to gene flow with many fewer investigations into source/sink dynamics, detection of movement corridors, or understanding functional connectivity. Perhaps the main reason so many investigations were limited to making descriptive evaluations was that there was such a large diversity of species-specific results (from Storfer et al 2010):
To summarize, the effects of a wide variety of landscape variables on population genetic structure have been considered. Several landscape variables such as elevation, ridgelines and topographic relief limited gene flow in several, but not all terrestrial species. In aquatic species, drainage structure, slope, elevation and temperature were important explanatory variables. Anthropogenic features, such as deforestation, agricultural development, damming and other types of waterway manipulation decreased gene flow or affected movement pathways in some species, but had little to no effect on others. Some features, such as rivers and post-fire regenerated habitats facilitated gene flow, contrary to expectations. Taken together, this wide variety of studies suggest that, while there are some generalities, effects of landscape variables and habitat fragmentation vary among species, highlighting the need for species-specific studies.
As methodology from both genetic and landscape ecology perspectives continued to improve, other questions were rolled in under the landscape genetics terminology as well. Multiple earlier reviews mention the potential of studying the effects of landscape on adaptive genomic variation (Manel et al 2003, Holderegger and Wagner 2008), and these questions begin to surface as sequencing technology reduced the difficulty in identifying potentially adaptive genes (Manel et al 2010, Manel and Holderegger 2013). Community Landscape Genomics (Hand et al 2015) builds upon all of these frameworks by expanding the suite of explanatory variables for neutral/adaptive variation to both ecological and landscape parameters.
The Data
What a landscape genetics dataset looks like has changed extensively in the last twelve years, but this change does not differ too drastically from other field that use molecular data. Manel et al (2003) predicted the rise of SNPs as common markers, but Storfer et al still reported a majority of studies using AFLP and microsatellite data in 2010:
As we look to the future, recent technological advances will lead to the availability of vast amounts of molecular and even complete genomic data available for landscape genetics studies (Holderegger & Wagner 2008). Landscape geneticists should begin to consider how to incorporate new types of genetic data into landscape genetic analyses, and spatial analysis of genomic data in particular will likely require new methods and approaches (see Manel et al. 2010).
It isn’t until recently (Hand et al 2015) that reviewers have considered next-generation sequencing as the new standard of genetic data collection, and less “future direction” emphasis placed on our ability to generate higher resolution genetic data. Instead, the emphasis of the future is on the analytical models to make sense of the growing piles of information (from Petren 2013):
One might suspect that the driving force behind evolutionary landscape genetics would be the technical advances of modern genomics. This is part of the story, but a closer look reveals that the development of statistical and modeling approaches is equally responsible.
The Analyses (or, where did all the Mantels go?)
What I think is the most fascinating, and probably hardest to understand, facet of landscape genetics is the plethora of statistical methodologies that have been used and continue to be developed.
Because of its interdisciplinary nature, there is no roadmap of how to conduct a landscape genetics study. As a result, researchers from several different backgrounds face many decisions regarding sampling design, choice of molecular markers, and data analysis methods. (from Storfer et al 2010)
Some of the factors that Manel et al (2003) found limiting are certainly much improved in 2015: detecting adaptive loci is easier, simulation studies are more prevalent, and sample sizes are up. However, there is still no best way to analyze a landscape genetics/genomics data set.
In 2009, there were two particularly helpful resources for understanding the options available and best practices. In the first, Guillot et al (2009) approached the gap in statistical techniques from a molecular ecology vantage point, suggesting ordination tools for exploratory analyses (see Jombart et al 2009), a careful consideration of your coarse population structure before looking at isolation by distance measurements, and Bayesian clustering methods that incorporate spatial information (such as GENELAND and GENECLUST). However, these techniques can also come with the potential cost of less clarity of interpretation, especially when producing synthetic variables.

Storfer et al (2010) showing the percentage of landscape genetics studies that use certain analysis techniques.
The second, by Balkenhol et al (2009b), provided a comparison of eleven different analysis methods across five common genetic/landscape scenarios (i.e. landscape boundaries, IBD only, landscape resistance). The most concerning result of these comparisons was the high type-I error rate produced by Mantel tests, which until then were the most common analysis technique (Storfer et al 2010). The suggestions of Balkenhol et al (2009b) reinforced the need for non-linear, multivariate techniques (canonical correspondence analysis, generalized dissimilarity modeling). However, there is still no best-suited general approach:
Considering the highly complex nature of landscape genetics and the large variety of landscape genetic research questions, it is unlikely that a single method will ever suit all research needs. Using multiple methods to analyze the same data will lead to increased certainty of inferences, and ensures that conclusions drawn from land- scape genetic analyses are valid.
In 2015, Mantel tests have largely disappeared in favor of some of the methods suggested by these previous reviews. Even newer statistical methodologies that include ecological, spatial, and genetic data (such as BEDASSLE) are now available to provide access to the explanatory power of additional ecological parameters.
In the long term, landscape genetics needs to find ways to measure gene flow directly from raw data (e.g., allele frequencies) and then use diverse types of multiple regression (in the case of a single response variable) or canonical redundancy analysis (for multiple response variables) when investigating gene flow in response to landscape configuration and composition. (from Manel and Holderegger 2013)
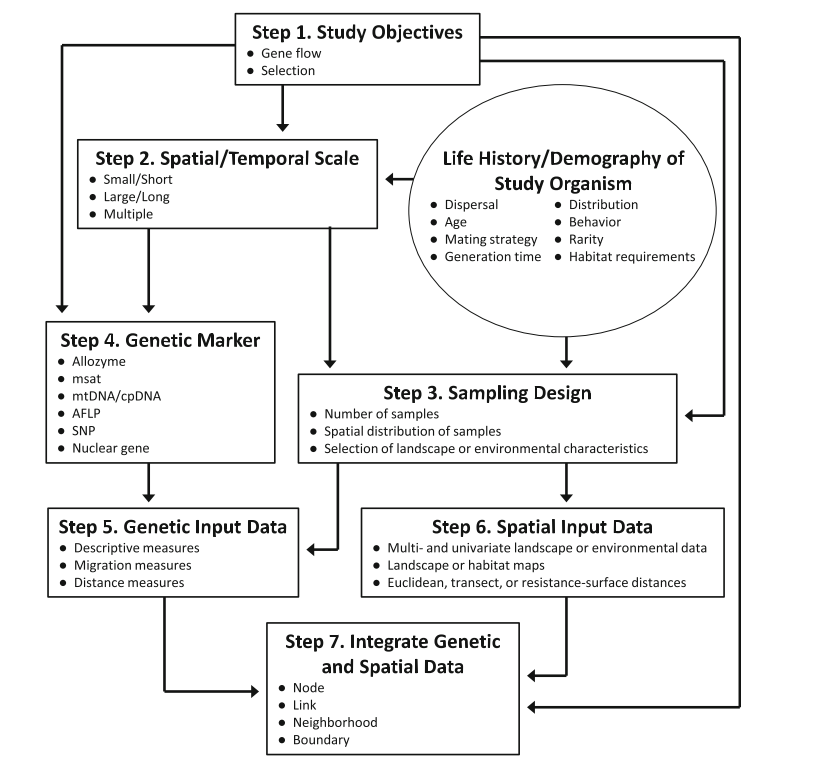
Hall and Beissinger (2014) provide a comprehensive and practical approach to study design
Moving forward the forward directions
Across all of these reviews, one common plea from authors has been to use knowledge based on organismal natural history to ask specific hypothesis. For example, Balkenhol et al (2009a), identified four challenges for the future of landscape genetics, three of which could be improved by simply testing a priori hypotheses that are driven by biological knowledge (spatial and temporal scale, analytical limitations, limited scope). The fourth challenge (lack of interdisciplinary communication) is something that has surely been improved by numerous workshops, symposia, and cross-journal special issues over the past twelve years.
Buried within multiple reviews of methodology, there has always been the suggestion of stepping back and being specific with your questions. Perhaps the race to use these methods to provide generalizable mechanisms has been premature, especially when considering the most often answer to any landscape genetics question is “it depends”.
Literature Cited
Balkenhol N, Gugerli F, Cushman SA et al. (2009a) Identifying future research needs inlandscape genetics: where to from here? Landscape Ecology, 24, 455–463.
Balkenhol N, Waits LP, Dezzani RJ (2009b) Statistical approaches in landscape genetics: An evaluation of methods for linking landscape and genetic data. Ecography, 32, 818–830.
Guillot G, Leblois R, Coulon A, Frantz AC (2009) Statistical methods in spatial genetics. Molecular Ecology, 18, 4734–4756.
Hall L a., Beissinger SR (2014) A practical toolbox for design and analysis of landscape genetics studies. Landscape Ecology, 29, 1487–1504.
Hand BK, Lowe WH, Kovach RP, Muhlfeld CC, Luikart G (2015) Landscape community genomics : understanding eco-evolutionary processes in complex environments. Trends in Ecology & Evolution, 1–8.
Holderegger R, Wagner HH (2008) Landscape Genetics. BioScience, 58, 199.
Jombart T, Pontier D, Dufour A-B (2009) Genetic markers in the playground of multivariate analysis. Heredity, 102, 330–41.
Manel S, Holderegger R (2013) Ten years of landscape genetics. Trends in Ecology and Evolution, 1–8.
Manel S, Schwartz MK, Luikart G, Taberlet P (2003) Landscape genetics: combining landscape ecology and population genetics. Trends in Ecology and Evolution, 18, 189–197.
Petren K (2013) The evolution of landscape genetics. Evolution, 67, 3383–3385.
Storfer A, Murphy MA, Evans JS et al. (2007) Putting the “landscape” in landscape genetics. Heredity, 98, 128–142.
Storfer A, Murphy MA, Spear SF, Holderegger R, Waits LP (2010) Landscape genetics: where are we now? Molecular Ecology, 3496–3514.